Genetik Kontrol ve Epigenetik Düzenleme
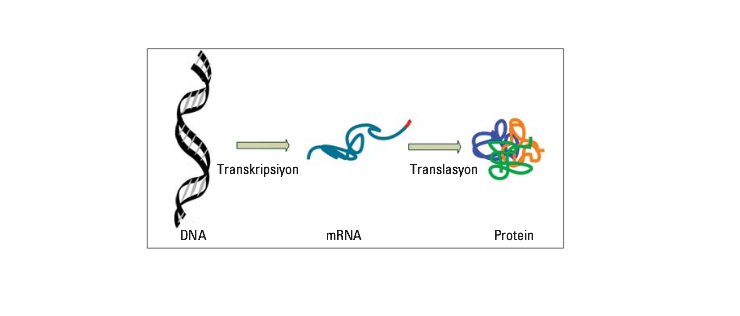
Francis Crick’in 1958’de ilk olarak sözünü ettiği moleküler biyolojinin temel dogması, nükleik asitle kodlanan bilginin mRNA üzerinden proteine translasyonunun olduğunu, bunun da tersinir bir ifllev olmadığını iddia eder (Şekil). Oysa 1980’lerden sonra bulunan moleküler kanıtlar bunun zaman zaman tersinin de söz konusu olduğunu, sekans değiflikliği ile sonlanmasa bile çevresel faktörlerin DNA transkripsiyonunu, hem nicel hem de nitel olarak değifltirebildiğini ortaya koymaktadır.
Epigenetik düzenlemeler birçok farklı mekanizmayla geliflmektedir (Tablo 1). Bunlardan en iyi bilineni DNA-metilasyonudur. Bu, yaklaflık 1 kb uzunluğundaki CpG adaları adı verilen, %50’den fazla nükleotidin sitozin ve guaninden olufltuğu promotor bölgelerindeki sitozinin (C) metilasyonudur. DNA-metiltransferazlarca (DNMT) yapılan bu düzenleme ile genin transkripsiyonu susturulmaktadır. Bu enzim mitozla ortaya çıkan yeni DNA’nın metillenmesi ve epigenetik bilginin korunmasını sağlamaktadır. Bu değiflikliğin mitozla aktarılabilmesi, çevresel faktörlerin uzun süreli etkilerini açıklamak açısından önemlidir.
Bazı hastalıklarda promotor bölgelerinin nasıl olup da uygunsuz flekilde hipermetile edildiği bilinmezliğini korumaktadır. Bu konuda değiflik hipotezler mevcuttur. Bunlardan biri de DNMT’lerin gereğinden uzun süre ortamda bulunmasıdır. Her enzim gibi bir ömrü olan DNMT’in ortamda daha uzun süre yıkılmadan kalan varyantları, DNA’nın hipermetilasyonunun sebebi olabilir. Bu ayrıca, demetilazların yetersizliği ve ikincil olarak DNA metillenmesine neden olan histon metilazların hiperaktivitesi sonucu da oluflabilmektedir.
Metillenmifl sitozin (5-MeC) deaminasyonla timine (T) kolaylıkla dönebilmektedir. Bu nedenle DNA’nın metilasyonu mutasyonel sıcak noktalar yaratarak genomik mutasyonlara da zemin hazırlamaktadır (5) .
DNA’nın metilasyonu ile ortaya çıkan kromatinin tekrar modellenmesinin bilinen en iyi örneği, adını Mary Lyon’dan alan X kromozomunun rastlantısal inaktivasyonudur (Lyon hipotezi). Aslında X kromozomundaki genlerin çoğu cinsiyet ile iliflkili genler değildir. Bu nedenle erkek ve kadınlarda eflit miktarda translasyonunun olmasının sağlanması gerekir. Bundan ötürü difli hücrelerde fazla olan bir X kromozomu (X inaktif) DNA metilasyonu ile inaktive edilip heterokromatin haline getirilerek, nükleusda Barr cisimciği olarak korunmaktadır. Herfleye rağmen X inaktif ‘in kodladığı genlerin yaklaflık %30’unun transkripsiyonunun olduğu düflünülmektedir. X kromozomundaki yapısal değifliklik bu genlerin inaktivasyonunda sorunlar çıkararak, susturulması gereken genlerin eksprese olması sonucunu doğurabilir. Bu da ilgili genin fazla ekspresyonuyla sonuçlanarak hastalık sebebi olabilir. X kromozomundaki kırılmaya uygun sıcak bölgelerin çokluğu (6) göz önüne alınırsa bazı hastalıkların (X inaktif )’den kaynaklanmasının ihtimal dahilinde olduğunu düşünmek gerekir.
Kaynak:
Nöropsikiyatri Arflivi 2008; 45 Özel Sayı: 15-20
Archives of Neuropsychiatry 2008; 45 Supplement: 15-20
BİYOLOJİ ÖDEV YARDIM
-

Mercanlar ve Mercan resifleri hakkında bilgi
-
Kulak Nedir? Kulağın Yapısı ve Görevleri Nelerdir?
-
Göz nedir ? Gözün görevleri nelerdir ? Canlılarda göz ve görme organı
-
Boğaz nedir ? Boğazın kısımları nelerdir ?
-
Omurga, columna vertebralis nedir ? Görevleri nelerdir ?
-
Doğal gübreler nelerdir
-
Kimyasal (yapay) gübreler nelerdir
-
Kortizol Nedir
-
Semantik Nedir ?
-
Karasal Ve Sucul Biyomların Özellikleri Nelerdir ?
-
Kaç çeşit biyom vardır
-
Bitki Ve Hayvanların Yeryüzündeki Dağılımını Etkileyen Faktörler Nelerdir?
-
Bitkisel dokular hakkında bilgi
-
Ekosistemde besin zinciri ve besin ağının önemi nedir ?
-
Genetik Algoritmalar
