Bakterilerde Homolog rekombinasyon
Homolog rekombinasyon bakterilerde önemli bir DNA tamir sürecidir. Bakteri topluluklarında genetik çeşitlilik oluşturmakta da önemlidir, ama bu süreç ökaryotik genomlarıa çeşitlilik getiren mayotik rekombinasyondan büyük farklılık gösterir. Homolog rekombinasyon Escherichia coli adlı bakteride ayrıntılı bir şekilde çalışılmıştır.[2] Bu yolağın iki versiyonu vardır, bunlar, çift iplikli kırıkların tamirini sağlayan RecBCD yolağı ve tek iplikli kırıkların tamirini sağlayan RecF yolağıdır.[3] RecBCD yolağı, bozulmuş veya duraksamış DNA ikileşme çatalını yeniden harekete geçirmeye, ayrıca transpozonlarda gen ifadesinin düzenlenmesinde kullanılır.
RecBCD yolağı
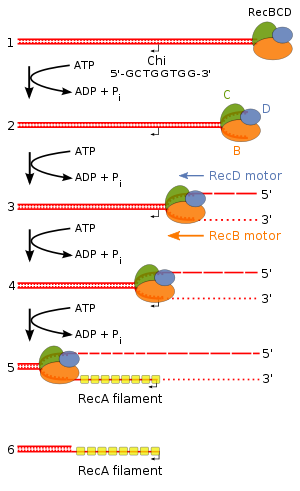
RecBCD yolağı, bakterilerde çift iplikli DNA kırıklarını onarmakta kullanılan ana rekombinasyon yolağıdır. Çift iplikli DNA'daki bir kırığın küt veya yaklaşık küt ucuna bağlanarak rekombinasyonu başlatır. RecBCD DNA'ya bağlanınca, RecB ve RecD altbirimleri, ATP hidrolizi ile yürüyen helikaz etkinliği sayesinde, birbirine sarılı DNA ikilisini çözmeye başlar. Bu iki altbirim sonra ayrılmış olan iplikleri endonükleotik olarak keser: RecB, 3' ucu, RecD'nin 5' ucu kesmesinden daha sık olarak keser. DNA'nın açılması ve kesilmesi RecBCD'nin Kay konumu (İng. Chi site) olarak adlandırılan spesifik bir nükleotit dizisine (5'-GCTGGTGG-3' ) varana kadar devam eder.[5]
Kay konumuna varınca RecBCD enziminin etkinliğinde dramatik bir değişiklik meydana gelir.[6] RecBCD birkaç saniye kadar duralar, sonra ilk hızının yarısı bir hızla ilerlemeye devam eder. RecD kesme hızını artırır ve 5' ipliği daha parçalanmış bir hale getirir, bu arada RecB altbirimi kesme etkinliğini durdurur. Kay konumunun tanınması RecBCD enzimini değiştirir, öyleki 3'uzantıya birçok RecA proteinleri yüklemeye başlar. Meydana gelen, RecA kaplı nükleoprotein filaman, homolog kromozomda benzer diziler aramaya başlar. Başka bir DNA molekülünde benzer bir dizi bulunca RecA'lı iplik, iplik işgali denen bir süreç ile bu homolog DNA ikilisinin içine girer.[5]
İşgalci 3' çıkıntı içine girdiği DNA ikilisindeki iki iplikten birinin yerini alır. Sonuç, Holliday bağlantısı olarak adlandırılan, X-harfi şeklinde bir yapıdır. RuvA Holliday bağlantısına bağlanır ve RuvB'yi seferber eder. Holiday birleşiminin DNA üzerinde yürümesi dal ilerlemesi olarak adlandırılır ve heksamerik bir ATPaz olan RuvB tarafından katalizlenir. RuvC zayıf spesifiteli bir endonükleazdır, junction ayrışmadan önce onun bir müktar ilerlemesine izin verir.[7]
Rekombinasyon iki tip ürün verir, bunlar Holliday birleşiminin nasıl ayrıştığına bağlıdır bunlara uç birleştirme (İng. splice) veya yama (İng. patch) denir. Uç birleştirme ürünleri yarım krosover ürünleridir, bunun sonucu genler karılır. Buna karşın yama ayrışması krosover olmayan ürünler meydana getirir (8)
RecF yolağı
Bakteriler DNA'daki tek iplikli kırıkları onarmak için homolog rekombinasyonun RecF yolağını (RecFOR yolağı olarak da adlandırılır) kullanırlar. RecBCD yolağı mutasyonlar sonucu çalışmaz hale gelince RecF yolağı çift iplikli kırıkların tamirinde onun yerini de alabilir. RecF yolağı hâlen RecBCD yolağından çok daha az anlaşılmış durumdadır. İki yolak da iplik işgali için RecA'ya gerek duyar, ayrıca dal ilerlerlesi (İng. branch migration) ve ayrışma evrelerinde benzeşirler.[9][10]
RecF yolağının başlangıcında, RecJ (tek iplikli DNA'yı 5' → 3i yönünde kesen bir eksonükleaz) DNA'daki tek iplikli bir kırıktaki 5' uca bağlanır, bu uçtan kırparak 3' doğrultuda ilerlemeye başlar. RecJ tek başına da çalışabilse de, tek iplik bağlayıcı protein (İngilizce single-strand binding protein veya SSBP) ve RecQ helikaz etkinliğinin mevcudiyeti 5' ucun ne kadar kırpıldığını (rezeksiyon miktarını) belirler. RecJ'nin 5' ucu kırpmasının ardından meydana gelen 3' çıkıntıya SSBP bağlanır.[10] Bu sayede tek iplikli 3' çıkıntının, komplementer diziler yüzünden kendi üzerine katlanmasının önüne geçilmiş olur.[11]
RecA'nın SSBP-kaplı 3' çıkıntıya yüklenmesi iki farklı yoldan olabilir, bunlardan biri RecFOR enzimini gerektirir, öbürü ise RecOR enzimini.[12] RecFOR yolağında, 3' uca sahip tek iplikli DNA'nın çift ipliklikçikli DNA ile birleştiği konumda RecFR kompleksi bağlanır. RecO sonra tek iplikli DNA üzerinden SSBP'nin yerini alır ama SSBP RecO'ya bağlı kalır. RecFOR sonra bu tek iplikli DNA/çift iplikli DNA birleşmesindeki (junction) içeri girinti 5' uca RecA'yı yükler. RecFR'de bulunan RecR altbirimi RecO ile etkileşerek RecFOR kompleksini oluşturur. Bunun sonucu RecR hem RecO'dan SSBP'yi ayrıştırır hem de 3' çıkıntıya RecA proteinleri yükler.[13]
RecOR tarafından RecA yüklemesi RecFOR yüklemesinden birkaç yönden farklıdır, özellikle moleküler etkileşim gereksinimleri ve ideal DNA substratı için.[12] RecFOR yolağında farklı olarak, RecOR yolağında RecO ile SSBP'nin C-ucu arasında etkileşim vardır. Ayrıca, RecOR yolağında 3' çıkıntıya RecA'yı bağlamak için tek iplikli DNA/çift iplikli DNA birleşmesine gerek yoktur, buna karşın verimli olması için RecFOR yolağında böyle bir gereksinim vardır. Bu yüzden recA yüklemesinde RecOR yolağı çoğu durumda RecFOR yolağından daha verimli çalışır.[12]
Dal ilerlemesi
İplik işgalinin hemen ardından, dal ilerlemesi olarak adlandırılan bir süreç ile, Holliday bağlantısı bağlanmış DNA üzerinde hareket eder. Holliday bağlantısının bu hareketi sırasında iki homolog DNA ikilileri arasında baz çiftleri birbiriyle yer değiştirir. Dal ilerlemesini katalizlemek için RuvA proteini Holliday bağlantısını önce tanır, sonra da ona bağlanır, ardından RuvB'yi de seferber ederek RuvAB kompleksini oluşturur. RuvB molekülü halkasal, altıgen şekillidir; iki RuvB molekülü Holliday bağlantısının iki tarafında yüklenirler ve dal ilerlemesine güç verecek birer pompa gibi çalışırlar. İki RuvA tetrameri Holliday bağlantısının kare-şekilli merkezinde yer alır ve DNA'yı sandviç gibi aralarına alırlar. Her bir DNA ikilisinin iplikleri RuvA'nın yüzeyinde çözülürler, protein onları bir ikiliden öbürüne yönlendirir.[14][15]

Ayrışma
Rekombinasyonun ayrışma (rezolüsyon) aşamasında, iplik işgal sürecinde oluşan Holliday bağlantıları kesilir, böylece tekrar iki ayrı DNA molekülü meydana gelir. Bu kesme işlemi, RuvC ile etkileşen RuvAB kompleksi beraberce oluşturduğu RuvABC kompleksi tarafından yapılır. RuvC bir endonükleazdır, 5'-(A/T)TT(G/C)-3' dizisini keser; bu dejenere kod, DNA'da sıkça bulunur (yaklaşık her 64 nükleotitte bir).[15] Kesmeden önce, RuvC Holliday bağlantısındaki iki RuvA tetramerinden birini yerinden çıkarıp oradaki DNA'ya temas eder.[14] RuvC'nin DNA'yı nasıl kestiğine bağlı olarak rekombinasyon ya "uç birleştirme" (İng. splice) ya da "yama" (İng. patch) ürünleriyle sonuçlanır.[15] Uçbirleştirme ürünleri krosover ürünleridir, rekombinasyon noktasının iki yanındaki genetik malzeme farklı şekilde biraraya gelir (asortisman olur). Yama ürünleri, buna karşın, krosover olmayan ürünlerdir, genetik malzemenin yeni bir bileşimi olmaz, rekombinasyon ürününde sadece hibrit DNA'dan oluşan bir "yama" vardır.[16]
Genetik transferin kolaylaştırılması
Yatay gen transferi, bir organizmanın, yavrusu olmadığı başka bir organizmaya ait yabancı DNA'yı edinme sürecidir. Yatay gen transferinde verici organizmanın DNA'sının alıcı organizmanın genomuna entegre olmasında homolog rekombinasyon önemli bir yoludur. Homolog rekombinasyon alınan DNA'nın konak genoma yüksek derecede benzer olmasını gerektirir ve bu yüzden yatay gen transferi genelde benzer bakterilerle sınırlıdır.[17] Pekçok bakteri üzerinde yapılan çalışmalar göstermiştir ki, verici ve alıcı organizmaların DNA'ların dizi benzerliği arttıkça rekombinasyon sıklığı log-lineer bir azalma gösterir.[18][19][20]
Genetik
-

İnsanlarda Kaç Kromozom Vardır?
-
Sık görülen mikrodelesyon sendromları nelerdir?
-
Bilim insanları kromozomları nasıl inceler?
-
Arkea'da Kromozomlar ve DNA Replikasyonu
-
DNA Onarım Mekanizmaları Nelerdir?
-
DNA hasarına neden olan etkenler nelerdir?
-
XYY Süper Erkek Sendromu - JACOB’S, Sendromu
-
Bitki doku kültürü çalışmaları ile haploid bitkiler elde edilebilir
-
Gram pozitif bakterilerden genomik DNA izolasyon protokolü
-
E. coli bakterisinden genomik DNA izolasyon protokolü
-
DNA’nın Keşfi
-
İnsan Genom Projesi Nedir ? Amaçları Nelerdir ?
-
Genomik mikrodizilimlerle ikilenme teşhisi yöntemi
-
Gen duplikasyonu ve amplifikasyonu nedir?
-
DNA ile RNA Arasndaki Farklar ve Benzerlikler Nelerdir
